

(本文由 Nautilus 授权转载,撰文/Sean H.Rice)性无处不在。绝大多数的动物和植物都至少会在某一阶段有性生殖。有的物种,比如人类,只能有性生殖。不过,弄清楚为什么有性生殖如此普遍一直是演化生物学家的老大难——因为,作为一种生殖策略,有性生殖似乎纯属浪费。
性有两个问题。一是它会打乱你的基因组合:你成功地活到了成年,这一点就足以表明你的基因现状相当适应环境了;而如果现在把你的基因打乱、和别人的基因混在一起,能不能用尚且是未知数,更别提会不会有改进了。二是它需要两个人才能完成,每人只提供一半基因:如果一位雌性决定改用无性繁殖,把自己的基因完整原样传给后代,她就能传递双倍的基因。这被称为“减数分裂代价”。总体而言,克隆自己似乎才应该是更好的选择啊?
 绘图:Yoko Shimizu
绘图:Yoko Shimizu
诚然,有性生殖和基因重组产生新变种的速度比单靠突变来得快,这对生物体适应不可预测的环境可能有所帮助。但是,这足以抵消有性生殖给个体造成的短期损失吗?从20世纪中叶一直到20世纪末,这个答案似乎一直是“不能”。很多种群遗传学家建立了模型研究这个问题。在模型中,一个“修饰基因”决定包含它的基因组会不会经历重组(修饰基因的一个等位基因允许重组;另一个则阻止重组)。大多数情况下,这些模型的重组频率都有所下降并最终消失。此外,有性生殖和基因重组往往降低了适应性演化的速度。约翰•梅纳德•史密斯(John Maynard Smith)总结了这些模型,得出的结论是:虽然重组可能在一个不可预测的环境中受青睐,但这个环境“不可预测的方式”必须“非常特殊,而且有些难以置信”。
事实上,这些模型含有一个简化了的假设。它虽然看似无奇,实则隐藏了一些关键过程,这些过程令有性生殖更可能成为适应性特征。这个假设就是:种群是无限大的。在含有突变的无限大的种群当中,一切等位基因的一切可能组合都永远存在。只要环境发生变化,那个已经存在的最优基因型的频率就会立即上升——而重组则成了阻碍。
但是,在有限的种群中,等位基因的所有组合不是随时都有。此外,在基因漂变(参见“理解漂变”)的影响下,现存的组合中总有一些不太适应环境。近年来,研究人员越来越多地将注意力放在建立有限种群在不可预测的环境中的模型上面。在这种情况下,等位基因的绝大多数可能组合起初并不存在。有性生殖和基因重组将最初出现在不同个体里的等位基因汇集在一起,创造有利的组合。进一步理解生物根据环境进行无性或有性生殖后会发现,在一个不可预知的环境中通过基因重组产生自适应变异是维持有性生殖这一繁殖策略的合理解释。
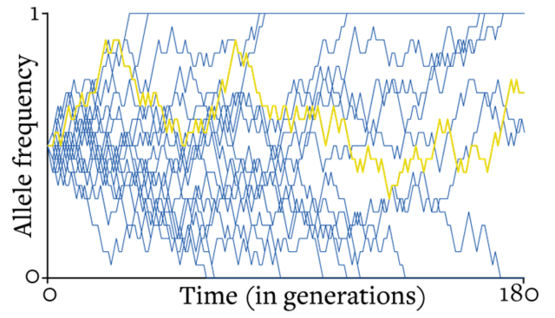
小贴士:理解漂变
要弄清楚基因漂变对演化的影响,以及我们能对它做出怎样的预测,需要明白两件事情。第一:随机漂移也会漂出“方向”来。具体讲,随着时间的推移,随机漂移会离起点越来越远。我们无法预测系统朝哪个方向漂,但可以预测在某一时刻系统所到之处的位置分布,而随着时间流逝,这个分布会逐渐向外散开。第二:一旦某个等位基因在种群中固定或灭绝,它就会停止漂移。学术上,我们说频率为0(等位基因灭绝)和频率为1(等位基因固定)为“吸收态”。下图显示了这两种情况的后果。图中画出了19次基因的随机走动,每一步都代表了随着时间的推移,等位基因在不同种群中的出现频率。其中一个种群用黄色高亮,让你能看到单次随机行走是啥样子的。经过180代以后,只剩下3个不同的随机行走路线还在进行,剩下的全都达到了0或1的吸收态、不再改变。不过,任何有限种群抵达0或1都是迟早的事儿。
就算是在大种群里,漂变也是很重要的。哪怕是对于一个任意大的种群来说,某个中等适应程度的突变也不见得一定会固定下来。比方说吧,一个突变如果能让有效后代数量增加10%,那么它也只有18%的概率能固定下来(82%的情况下它都会丢失),即使在个体数量高达千亿的种群里也是如此。关键原因是,当一个等位基因刚出现的时候,它只有一个或几个副本,而它们因为数量太少,命运里的偶然因素太大了,这和有多少别的个体无关。换言之,就算特别巨大的系统,它们的表现也和假想的无限大的系统有重大差异(参见“理解无限”)。
这个例子说明了我们在考虑浪费和效率时常犯的错误。我们往往直观地以为世界是无限和确定的。当面对有限的、有随机性的世界时,我们常常不加批判地将现实视为“最佳”世界的粗略近似版;要不是一些恼人的瑕疵,我们就会有一个完美的世界。
这一柏拉图本质论的现代余烬——要获得真正的洞见就应该研究理想世界的完美形态——有时令我们无法认清有限系统并不只是无限系统的模糊近似(参见“理解无限大”)。相反,有限系统的行为方式有这本质上的不同,这些差异对只考虑理想无限世界的人来说想都想不到。
小贴士:理解无限
人们通常以为无限大是个很大很大的数目。其实不是的。严格说,这篇文章里讲的大多数概念在真正无限大的种群中都是没有意义的。科学家说对“无限大”种群建模时,他们真正在做的是研究等式在种群数量越变越大趋于极限时的表现。无限大并不是一个数。要是真将种群数量视为无限大,就无法确定等位基因的出现频率,因为不同的数字除以无限大,得到的结果是无法区分的。
在进化生物学中,这意味着研究有限种群会发现全新的过程。在概率演化过程中被研究得最透彻的是遗传漂变——小群体内基因频率的随机波动。科学家通常将漂移想象为“随机游走”的情形,每一代中,变体的频率可能向上一步(走向固定)、向下一步(走向灭绝),或者保持不动(参见“理解漂变”)。向上或向下一步的大小与种群数量成反比。在大种群中一步很小,而小种群里这一步就很大(这并不表示漂移在大种群中不重要)。
因此做出适应性演化涉及到多次尝试,其中有许多都是不成功的,甚至在错误的方向上迈出的步子。有限种群的适应性演化实际上需要常常出错的探索。因为相似的基因组可以产生区别很大的表型,其适应度大相径庭。打个比方,一座山有高低不同的很多山峰,登山者想要攀登最高峰,但没有办法看到全景。最简单的办法是沿着上坡走,但很可能会被困在一座较低的山头。解决这个问题的办法就是偶尔走下坡——时不时接受比当前更糟的解决方案。
事实上,这正是包括遗传算法和模拟退火在内的很多等现代算法采用的思路。这些算法找到复杂问题最优解的方法,即是在有些情况下接受比当前情况更糟糕的解决方案。只要让算法对较差解决方案的接受度逐渐降低(在我们越爬越高的时候也越来越少选择下坡路),这种算法最终会趋近最佳值。一台拥有无限的处理能力和内存的计算机不需要使用遗传算法解决复杂问题。它可以并排比较所有可能的解,就像拥有无限多个体的种群不需要依靠有性繁殖以适应环境变化那样。但是,资源有限的计算机必须不时后退一步,才能再向前迈出两步。
我们可以在艺术、科学等人类创作中发现相同的有限性。科学的目的是增加我们对宇宙的客观特质的认识和了解。艺术的目的是探讨和交流个人体验的主观感受。因为宇宙和我们在其中的生活极端复杂,科学家和艺术家的很多探索都遭遇死角是并不奇怪的。如果一个科学家或艺术家无所不知,他就不用把时间浪费在没有解的方程、没有提供有用信息的实验或不成熟的隐喻之中。
我们之所以认为不成功的变种纯粹是浪费,是因为我们默认系统是无限大和理想化的。在现实世界这一充满复杂难题的有限系统中,不断试错再加上某种形式的选择,不仅是可行的做法,更是意外高效的选择——不管是对于一个鸡蛋还是对于一个抽象表现主义艺术家,都是如此。
肖恩•赖斯(Sean Rice)是德克萨斯技术大学的生物学教授,著有《进化理论:数学和概念基础》(Evolutionary Theory: Mathematical and Conceptual Foundations)。
本文由 Nautilus 授权果壳网(guokr.com)编译发表,严禁转载。
-
编译自:Evolution in a Finite World
-
原创人员:撰文/ Sean H. Rice;绘图/ Yoko Shimizu