
相信大家都熟悉《旅行青蛙》这款“佛系”小游戏吧!有很多小伙伴每天乐此不疲的在游戏中收三叶草、买点心、买帐篷,盼着呱儿子能多寄些漂亮的明信片回来。
游戏中的呱儿子无忧无虑活泼可爱,殊不知,现实世界中“旅行”的蛙,正在给它们的野生同胞带来一场可怕的灾难!
 吓得我赶紧打开游戏看看儿子回来没……(图片来源:《旅行青蛙》截图)
吓得我赶紧打开游戏看看儿子回来没……(图片来源:《旅行青蛙》截图)
全球蛙类“神秘失踪”事件
蛙作为一种古老的两栖动物,在地球上已经生活了1.9亿年。早在《旅行青蛙》出现之前,它们的足迹就遍布世界各地了——可以说,只要有水塘的地方,就有蛙的身影。
然而,生物学家通过近几十年来对两栖动物的野外调查发现,全球正在发生多起呱儿子“神秘失踪”事件——在20世纪70年代,美国、波多黎各和澳大利亚等地区记录到蛙类数量迅速下降的现象;80年代后期,在哥斯达黎加某地,40%的蛙类在短时间内神秘消失;厄瓜多尔和委内瑞拉等地也发现了蛙类突然失踪的现象……[1]
 研究者正在调查死亡的蛙,图片来源:Science
研究者正在调查死亡的蛙,图片来源:Science
全球的蛙类数量似乎都在突然之间急速下降。
开始,这些发现还受到很多研究者的质疑,毕竟两栖动物种群波动本来就很大。但随后更多数据显示,这些神秘失踪事件造成的波动幅度明显要比正常的波动更为广泛和严重[1]。
随着蛙类失踪事件的增多,更多的研究者们意识到事情确实没有想象的那么简单——蛙类数量减少甚至灭绝并非偶然事件。国际世界自然保护联盟(IUCN)调查指出,全球蛙类的下降幅度和严重性已经远高于其它脊椎动物[2] 。
然而究竟是什么导致了呱儿子们集体消失的呢?最初,研究者们也一直没有搞清楚,直到1998年,呱儿子“职业杀手”——蛙壶菌(Batrachochytrium dendrobatidis,简称Bd)被科学家缉拿归案,“蛙类失踪之谜”终于被破解[3]。
壶菌为何方神圣,如何变成呱儿子杀手?
蛙壶菌是一种真菌,隶属于壶菌门(Chytridiomycota)。这类真菌的独特性在于它们具有能动的、鞭毛状的游动孢子,不但在水生环境和土壤中能够自由生活,还能寄生在藻类、无脊椎动物、真菌和植物中,并感染包括蛙类在内的各种两栖动物。蛙壶菌是壶菌家族中对蛙类危害最为严重的一种。
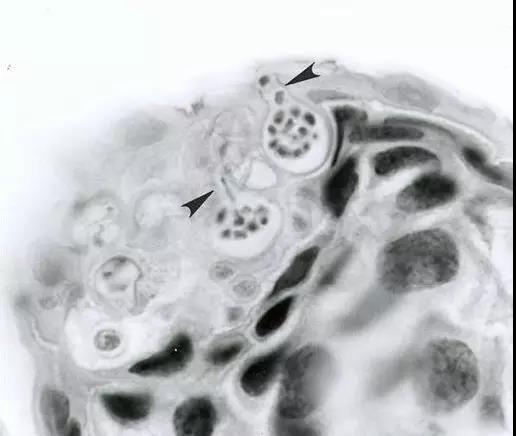 寄生在蛙皮肤上的蛙壶菌(Batrachochytrium)——箭头指向的是蛙壶菌孢子囊在产生新的孢子(图片来源:Wikipedia)
寄生在蛙皮肤上的蛙壶菌(Batrachochytrium)——箭头指向的是蛙壶菌孢子囊在产生新的孢子(图片来源:Wikipedia)
蛙壶菌“谋杀”蛙类的手段非常残忍——它们会在蛙表皮角质细胞内生长和感染,导致蛙的表皮增生、大量蜕皮,同时抑制蛙体内电解质转运,影响心脏功能,最终蛙因心跳停止而死亡[3,4]。
蛙壶菌对不同种类蛙的危害存在差异——对于一些敏感的蛙类(如绿雨滨蛙),感染壶菌后的死亡率甚至高达100%![5]目前,全球已检测的约1300种两栖类就有将近700种感染蛙壶菌,感染比例已超过50%。此外,由于蛙壶菌喜湿怕热(一般超过30度便很难存活),在气温较低的山区溪流中生活的蛙类比其他地区的更易感染[6] 。
 被蛙壶菌“谋杀”的产婆蟾(皮肤已大面积脱落)图片来源[7]
被蛙壶菌“谋杀”的产婆蟾(皮肤已大面积脱落)图片来源[7]
为什么呱儿子成了主要受害者?
呱儿子的生活发育史可以说为蛙壶菌提供了“优越”的侵染条件。
由于两栖动物的卵是非羊膜卵,没有卵壳,无法在空气中维持形态和防止水分蒸发,因此只能在水中产卵。呱儿子的儿子——蝌蚪,孵化后具有角质化的口盘,容易被水中的蛙壶菌侵入。而变态期蛙类自身的免疫力会降低,随着全身皮肤角质化程度增加,会导致变态期以及成体蛙大量感染壶菌。同时,蛙类的皮肤需要保持湿润才有辅助呼吸的功能——这一系列生活史特点正好被蛙壶菌所利用。
其他动物如水鸟、鱼也可能携带蛙壶菌但通常不被感染。偶尔也有一些例外,如一些蛙壶菌可导致斑马鱼的鳍或小龙虾的鳃受损或死亡,这些小龙虾也可以通过水体将蛙壶菌传染给呱儿子[8,9]。
蛙壶菌的“家谱调查”
最早,蛙壶菌被分为4个株系,即全球株系(BdGPL), 非洲株系(BdCAPE), 欧洲株系(BdCH)和巴西株系(BdBRAZIL)[10-12]。相比于3个地方性株系,全球株系分布最广,且大多毒力强,最可能导致两栖类种群下降或灭绝,因此成为重点研究对象。
生物学家们对于蛙壶菌全球株的来源问题一直争执不休,不同学者获得调查结果也差别极大。有研究者们推测全球菌株的祖先可能出现在100年前,也有研究说说可能出现在26000年前[10,11],发源地可能在非洲,北美,南美或东亚[13-16]。
直到近日,Hanlon等学者在《科学》(Science)期刊发表了一篇关于蛙壶菌起源的文章,进一步揭开蛙壶菌起源的神秘面纱[17]。
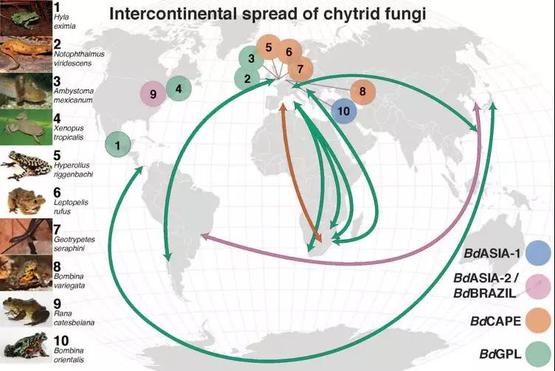 蛙壶菌的全球传播 | 图片来源[17]
蛙壶菌的全球传播 | 图片来源[17]
研究者为了确定蛙壶菌起源时间和起源地的问题,分离了来自亚洲、欧洲、北美、南美、非洲和澳大利亚的蛙壶菌,获得177种菌株的基因组,把它们与之前研究发表的基因组合并进行比较,生成具有全球代表性的蛙壶菌数据库。
通过对这个大型数据库的分析,研究者们发现了新的蛙壶菌BdASIA-1,并根据它的分布得出东亚地区是全球蛙壶菌多样性的中心区域。同时推测出蛙壶菌全球株系起源扩散的时间为50年前~120年前——这一时间段,恰好是全球贸易开始高速发展的时期!
人类竟成蛙壶菌传播“帮凶”
如此说来,造成蛙壶菌在全球传播的始作俑者,竟是人类。近一个世纪来,两栖动物的相关国际贸易——如实验动物、食用和宠物等需求,直接导致了蛙壶菌在世界范围内的传播。同时,人为活动也会无意识的带来两栖类的迁移以及蛙壶菌的传播,例如最近入侵马达加斯的亚洲蟾蜍,就是隐藏在采矿设备中逃避了海关检查,并最终扩散到马达加斯加。
 被人类端上餐桌的呱……图片来源:Science
被人类端上餐桌的呱……图片来源:Science
大量亚洲两栖动物在全球贸易中的运输,使蛙壶菌扩散的风险变得更高。比如从香港出口到美国的两栖动物中就发现了携带BdASIA-1蛙壶菌的东方铃蟾[18]。此外,其他重要的贸易种,如非洲爪蟾,北美牛蛙,墨西哥钝口螈等都是蛙壶菌携带者。看来,放任呱儿子去“旅行”,真的会加剧蛙壶菌在全球的传播。
 两栖动物国际贸易之一的东方铃蟾,图片来源:Science
两栖动物国际贸易之一的东方铃蟾,图片来源:Science
研究仍在继续
由于Hanlon等人新发现的BdASIA-1大多来自韩国的野生蛙,可能只代表了蛙壶菌原始菌株的一部分。亚洲其他地区急需进一步采样以完善数据库。
讲到这儿,你们可能会问道,中国的呱儿子们现在境况如何呢?其实,中国也有不少学者对蛙类野外样本、博物馆标本以及宠物市场做了大量调查,发现中国的两栖类表现为大范围低密度的感染,但幸运的是,目前还没有出现明显病症,也未发现大量死亡现象。
有学者推测,这可能是由于中国的本地蛙与蛙壶菌存在长期共同进化历史或者本地菌株的毒力较弱,也可能是野外调查的还不彻底。当然,上述推测尚需要进一步研究验证。另外,有学者指出中国也有很多适宜蛙壶菌生存的环境[19-21],因此还不能排除被感染的风险。为了呱儿子的健康,“防护措施”还是一定要加强的。
说到防护,肆虐全球的蛙壶菌也不是无药可治。例如,用广谱抗真菌剂伊曲康唑给病情较轻的呱儿子泡个澡就可以治好[22];可以通过高温灭活环境中的蛙壶菌来杜绝传染;还有学者们在探索对蛙进行益生菌治疗[23]。
不过,以上这些方案大多适用于人工饲养条件下的蛙,对于野外个体目前还没有行之有效的防治措施,只能选择迁地保护——赶在蛙壶菌侵袭之前,人为将蛙迁移到无壶菌的地区。
科学家们建议,除了保护蛙类栖息地,必须加强对两栖动物贸易的监管力度,防止蛙壶菌传播的进一步加剧。为了保护现存的野生蛙类,一定要给去“旅行”的蛙们做好“体检”!
 图片来源:科学人的“表情包女皇”——郅鸥供图
图片来源:科学人的“表情包女皇”——郅鸥供图
感谢Yuki小柒对本文章提供的修改帮助。
(编辑:明天)
参考文献:
- J. A. Pounds et al., Conserv. Biol. 11, 1307 (1997)
- S. N. Stuart et al, Science 306, 1783 (2004).
- L. Berger et al., Proc. Natl. Acad. Sci. 95, 9031–9036 (1998).
- J. Voyles et al., Science 326, 582-585(2009).
- L. Berger, Inter-Research, 68: 47–50 (2005)
- J. S. Piotrowski et al., Mycologia 96,9-15 (2004).
- M. C. Fisher et al., Annu. Rev. Microbiol. 63, 291–310 (2009).
- N. Liew et al., Nat. Commun. 8, 15048 (2017).
- T. A. McMahon et al., Proc. Natl. Acad. Sci. 110, 210-215 (2013).
- R. A. Farrer et al., Proc. Natl. Acad. Sci. U.S.A. 108, 18732–18736 (2011).
- E. B. Rosenblum et al., Proc. Natl. Acad. Sci. U.S.A. 110, 9385–9390 (2013).
- R. A. Farrer et al., PLOS Genet. 9, e1003703 (2013).
- C. Weldon, L. H. du Preez, A. D. Hyatt, R. Muller, R. Spears, Emerg. Infect. Dis. 10, 2100–2105 (2004).
- B. L. Talley, C. R. Muletz, V. T. Vredenburg, R. C. Fleischer, K. R. Lips, Biol. Conserv. 182, 254–261 (2015).
- D. Rodriguez, C. G. Becker, N. C. Pupin, C. F. B. Haddad, K. R. Zamudio, Mol. Ecol. 23, 774–787 (2014).
- A. Bataille et al., Mol. Ecol. 22, 4196–4209 (2013).
- Simon J. O’Hanlon et al., Science 360, 6389 (2018).
- J. E. Kolby et al., PLOS ONE 9, e 90750 (2014).
- W. Zhu et al., Dis Aquat Org. 118, 259–266 (2016).
- C. M. Bai et al., Diversity and Distributions. 18, 307-318 (2012).
- X. Liu et al., Proc Biol Sci. 280, 1753 (2012).
- N. Lubick, Nature 465, 680–681 (2010)
- R. N. Harris et al., ISME J. 3, 818–824 (2009).