
“The characters which naturalists consider as showing true affinity between any two or more species, are those which have been inherited from a common parent, all true classification being genealogical.”
-Charles Darwin, 1859:391
“博物学家们认为两个或两个以上的物种间那些表明真实亲缘关系的性状都是从共同祖先遗传下来的,一切真正意义的分类都是根据种系发生的。”
-查尔斯·达尔文,《物种起源》 1859: 391
波澜壮阔的系统学
“系统学”对很多人来说或许是个陌生的概念,但从人类开始认识自然,认识生命的多样性,系统学就是个绕不过去的核心:人们要认识世界万物,就要把它们分门别类、总结个性和共性,那么生命有机体的多样性是怎么形成的?如何去认识、发现、描述、解释这些生命的多样性?如何综合这些信息,去获得一个具预测性的分类系统?
这就是系统学。
在达尔文划时代的自然选择理论提出之后,系统学家们和演化研究逐渐走到了一起,广泛地说,现代系统学的根本目标是发现生命演化之树的所有分支,记录这些分支在演化过程中发生的变异,在最大程度上描述物种——这些分支的末端。因而,系统学是对地球上现存生物多样性及演化历史的研究。
对世间一切生物的分类系统都有两种:“自然”的,和“人为”的。对这两个词当然有许多的理解,但就目前系统学家们普遍共识是,能够反映演化历史的分类系统,才是“自然的”分类系统,除此之外皆为“人为”系统。举个例子,硬骨鱼、软骨鱼、鱼龙、鲸都有鳍或者鳍状的附肢,在一个“自然的”分类系统里,它们会被分到不同的小型类群:因为这极其相似的性状特征,是在不同的时期经由不同的路径演化而来的,它们是“趋同演化”的结果——并且它们之间最近的共同祖先,出现在不同的地质年代;而一个“人为的”分类系统,就可能因为“具鳍或者鳍状的附肢”这个特征,把它们归为同一个排开“不具鳍或者鳍状的附肢”(比如人类)的小类群——事实上我们当然知道,人和鲸的关系,比人或鲸和其它三者的关系都要近得多。
因此,泛泛地说,达尔文之前的系统学历史,是一部“人为”分类系统的历史,在植物界,包括林奈、Jussieu、Candolle、Benthem等植物学家的系统学工作,可归于此;在达尔文之后,追求“自然的”分类系统的进程如一股涓涓细流,最终波澜壮阔成为一片大海,也成了现代系统学的共识。
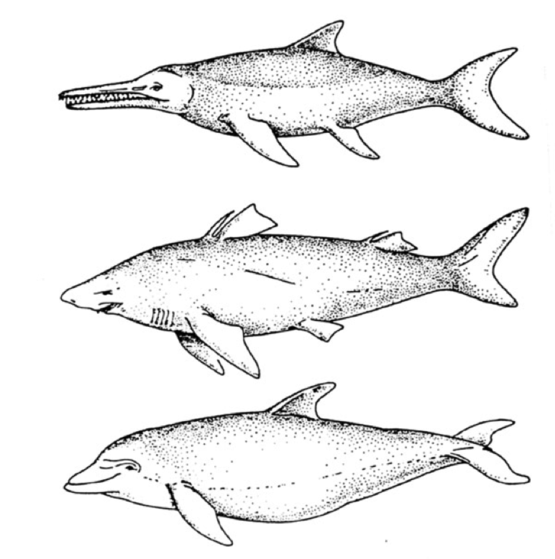
从上往下是鱼龙、鲨鱼和海豚,相似的外形和鳍状的附肢来自于趋同演化。图片来源:作者提供
在一个“人为”分类系统中,因为这些趋同演化的特征它们可能会被归到一类——虽然事实上它们关系很远。植物界也是如此。
因为“自然的”分类系统所基于的依据——物种水平以上的演化历史是惟一的,因此要获得一个“自然的”分类系统,第一步,我们要了解演化事件的顺序信息即系统发育(phylogeny),第二步,如果我们确切地知道了演化树的结构,剩下要解决的问题,就只剩下给演化树上的分支命名——很简单对不对?但事实上,直到人类能够大规模进行DNA测序之前,我们在“创造一个‘自然的’分类系统”这件事上的进展,一直被困在第一步。
我们都知道中心法则:在所有的生物体的细胞结构里,DNA自我复制,所包含的信息转录给RNA,RNA翻译为蛋白质,蛋白质和其它部分物质决定了生物体的一切,包括细胞结构、外观形态、生理行为……一旦包含有遗传信息、可以表达的DNA序列(基因)发生变化,则随之一切发生变化;在种群一级上,则是基因频率的变化——这就是演化。系统学家们一直在试图构建世间万物的系统发育结构,可用的数据就在这些演化所改变的东西上——DNA序列、RNA序列、蛋白质序列、细胞结构、微观形态数据、宏观形态数据、行为学数据……这些可以统称为“性状”。从某种意义上,任何一级的“性状”都对系统学家推断演化历史有重要意义,当然不同的是,分子序列可用的数据在数量上较之宏观形态要多得多。
在过去,尽管系统学家们努力还原演化历史,但苦于工具所限,从宏观结构到微观结构到化学成分,几乎止步于此。20世纪后期的科技成就包括计算机和大通量的DNA序列测序技术为系统学研究迎来了新的曙光,大量系统学的文章发表,这些进步也近乎颠覆性地改变了人类对现有生命类群的理解。
三张中文海报的诞生
高清版海报点击“这里”。
在有花被子植物领域,一群植物系统学家们组成了APG组(ANGIOSPERM PHYLOGENY Group, 被子植物系统发育组),共同讨论、推进、建立一个基于系统发育(即演化历史)的被子植物分类系统,除了最重要的单系原则(每一个被命名的分类阶元应该是单系的:即包含一个共同祖先和它所有的后代)之外,还应具有稳定性(尊重已有的类群名称,维持类群大小的稳定和适中,如无必要,不增加新的分类阶元或更改)和易用性(各被命名阶元和类群应具有明显的、可区别的形态特征)。第一版APG自1998年发表以来,被学术界广为采纳,英国皇家植物园和密苏里植物园均将各自标本馆中的馆藏顺序更改为按APG系统排列。然而因为众多历史原因,基于系统发育和演化的被子植物APG分类系统在国内还未得到广泛使用,给植物研究的国际化合作和交流带来了或大或小的困难。
去年,我在密苏里植物园的导师Peter Stevens教授告诉我,明年第四版的APG IV文章就要刊出了,并将其中的一些问题告知了我,这个消息令我非常兴奋。曾经协助他更新APG website的我一直非常关心APG系统在国内的普及情况,因此,当APG IV文章刊出后,我意识到这是一个契机。特别是在我在任教于2016年6月中国植物园联盟举办的第三届植物分类学培训班后,学员们纷纷反映希望我们能够更多地介绍当前国际通用的一些系统分类学概念,我觉得我有必要和几位同事合作,推动APG系统在国内的认知和普及。
我们讨论后认为,目前可以逐步地推开一些APG系统的周边。海报(poster)就是一种很好的形式,可以用直观的图形来表现所要告诉人们的信息,将信息高密度而又直观地集中在一张图上。我知道APG系统很早就有英文版海报,由德国海德堡大学(Heidelberg University)的Theodor C. H. Cole以及柏林自由大学(Freie Universität Berlin)的 Hartmut H. Hilger教授制作,在学术界产生了广泛的影响。我的工作就可以在他们的基础上进行了。
经过半个月的树形结构描画、翻译描述、翻译科目拉丁名、一些额外标记和取舍,三张中的第一张——APG系统中文海报诞生了。随后,我又花了三天时间翻译制作了同系列的“维管束植物系统发育树海报”(包含了所有的基于分子数据整理过的裸子植物、蕨类和石松类植物的系统发育关系),并与在中科院华南植物园工作的叶文博士合作翻译制作了“苔藓植物系统发育树海报”(包含了苔类、藓类和角苔类),如今包含所有“高等植物”系统发育信息的三张海报俱已完成。
虽然“高等植物”已经是个“旧”名词(“高等”无意义),在这里也不妨借用来指代所有陆生植物/有胚植物(Land plants/Embryophyte),包含苔类、藓类、角苔类、石松类、蕨类、种子植物这些单系类群,不包含绿藻、地衣以及其他藻类。
如何使用这三张海报?
这里的海报基于三张原版海报,Angiosperm Phylogeny Poster主要展示所有有花被子植物的系统发育,Tracheophyte Phylogeny Poster展示裸子植物、蕨类植物、石松类植物的系统发育,Bryophyte Phylogeny Poster展示苔类、藓类、角苔类植物的系统发育。
我们制作翻译的中文海报较原版发生了不少变动,比如APG IV原版海报中只包含了APG IV部分科,在此我根据2016年出版的APG IV文章已经将所有416个科填入,且各目以下重新按照Linear APG(APG线性)排列,方便查询和应用;苔藓植物海报部分原版有不少错误,我们在翻译过程中已经一一纠正。
我们提供多项下载选择:原版、纯中文版、科目拉丁名保留版,以及我们特别制作的中文-拉丁对照版,可以供对拉丁文不熟的同学对照记忆背诵。
关于如何使用这三张系统发育树海报查看信息,我进行一些增加梳理:
1、 查看目/科以上的系统发育和演化关系:
这个海报包含了所有高等植物类群的系统发育和演化关系,浏览整张图即可获得大量已知的系统发育信息,对整个高等植物有一个大略的了解。一些大类群上最显著的有趣信息比如:传统上的“苔藓植物”其实分为苔类、藓类、角苔类,“苔藓植物”是一个并系群,其中角苔类和所有维管束植物的关系,比它们和苔类或藓类之间的关系要近;传统上的“蕨类”包含真蕨类、木贼、石松和松叶蕨,但从这张海报你可以发现,松叶蕨可以包含在真蕨当中(和瓶尔小草目是姐妹群,有最近的共同祖先),木贼和真蕨关系紧密,而石松类和真蕨类关系较远——比真蕨类和所有种子植物的关系更远;传统上被子植物被分成单子叶植物和双子叶植物,而事实上,“胚具一片子叶”这一性状是从“胚具两片子叶”的类群中演化而来的,某些双子叶植物和单子叶植物之间的关系,比它们和另一些双子叶植物更近——前者被称为“真双子叶植物”。

几种常见的栽培植物的果实。左上为肉豆蔻科的肉豆蔻 Myristica fragrans,左下为无患子科的荔枝Litchi chinensis,右边为禾本科的小麦属 Triticum spp. 尽管肉豆蔻和荔枝在形态上有许多相似之处且都有两片子叶,但实际上小麦(单子叶植物)和荔枝(真双子叶植物)的关系比它们和肉豆蔻(木兰类分支)更近。图片来源:作者提供。
你可以在海报上找到你所感兴趣的两个以上类群,观察它们的拓扑结构,比如我想知道伞形科、蔷薇科和豆科之间的关系,我只需在图上找到这几个科,可见伞形科所在的伞形目,蔷薇科所在蔷薇目以及豆科所在的豆目的关系是((蔷薇目+豆目)伞形目),即蔷薇科和豆科之间的关系,比它们和伞形科之间的关系更近,那么蔷薇科和豆科较之它们和伞形科在距今更近的时间节点上有共同祖先,蔷薇科和豆科之间可能有更多的共同衍征。
2、 查看重要共同衍征:
在这张海报上,每个目以及目以上各分支的重要共同衍征都已经标注,这意味着:
你可以从海报上查看你想了解的目下一些重要的共同特征,如樟目Laurales:木本;叶对生;花有被丝托,常具退化雄蕊 花药常镊合状排列;心皮具1枚胚珠;胚大型。这些共衍征可以在樟目下各科中找到。

以从海报上查看你关注的类群的重要衍征。图片来源:作者提供。
你可以从海报上查看你所感兴趣的共衍征可能会出现在哪些类群上:比如“固氮”,我们可以看到这个共衍征出现在真双子叶植物-核心真双子叶植物-超蔷薇类分支-蔷薇类分支-豆类植物下的一个单系分支上,包含豆目、蔷薇目、葫芦目、壳斗目,那么如果我要寻找与细菌共生固氮的植物,就要在这几个目中寻找——在演化事实上也是如此,“与细菌共生固氮”这个共衍征,出现在以上四个目中的豆科、蔷薇科、桦木科、鼠李科、大麻科、秋海棠科、胡颓子科、杨梅科等科的部分植物中。
利用这些衍征做一些简单基本的检索。
 能够共生“固氮”的植物,出现在这些类群中。图片来源:作者提供。
能够共生“固氮”的植物,出现在这些类群中。图片来源:作者提供。
3、在被子植物系统发育海报中,查看APG IV的线性排列:
因为我已经将目下各科重新按照Linear classification of flowering plants(APG IV)排列,因此如果有标本馆(这是辰山植物标本馆正在做的工作)想将馆藏标本重新APG IV排列,那么可以参考这张海报。目下各科顺序为从上至下-从左至右,如下图所示。

海报中目下各科的LAPG顺序(用红色箭头表示)。图片来源:作者提供。
4、 查看中文-拉丁对照:
对于不熟悉拉丁科、目名的使用者,可以使用本海报进行对照记忆。
后记
三张植物系统发育树海报诞生了,受到了我没有想到的热烈欢迎,我将海报上传到“中国植物分类学家园”后又发了一条微博,三天的转载量达到了500多次,阅读量超过了25万——至少在我这里,这是个不错的结果。
当然我们不会止步于此,未来一年,我和辰山植物园的同事们还将推出一系列出版物,共同推进植物系统学的研究前沿在学界和爱好者界的认知,我期待有更好的结果。
毕竟演化,是生物学之光。
 自查尔斯达尔文1837年的手稿,在根本没有分子手段、甚至遗传学还没有诞生的十九世纪初,他清晰地写明了演化的共祖概念和树状结构。图片来源:作者提供。
自查尔斯达尔文1837年的手稿,在根本没有分子手段、甚至遗传学还没有诞生的十九世纪初,他清晰地写明了演化的共祖概念和树状结构。图片来源:作者提供。
部分海报细节展示:

APG4-poster主要展示所有有花被子植物的系统发育。
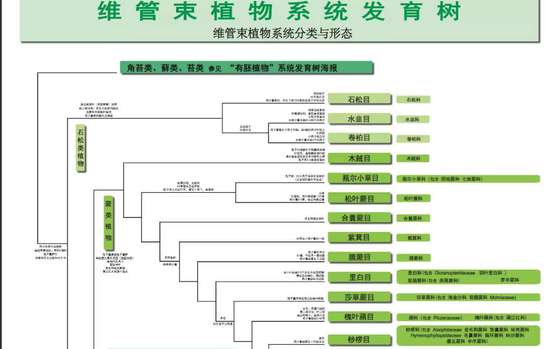
Tracheophyte Phylogeny Poster展示裸子植物、蕨类植物、石松类植物的系统发育
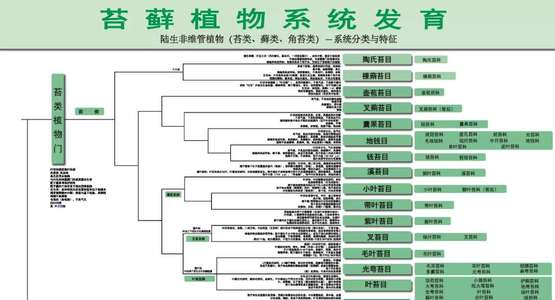
Bryophyte Phylogeny Poster展示苔类、藓类、角苔类植物的系统发育
点击“这里”下载完整清晰海报。(编辑:Sol_阳阳)